 |
Tweet |
GİRİŞ
Arkasında çiçekli güzel bahçesi olan bir evimiz olsun. Arka bahçemizin sınırının diğer tarafında ise komşumuzun evinin bahçesi uzanıyor olsun.
Bu sınırın tam üzerinde küçük birde çukur olduğunu varsayın.
Bir sabah komşunuzu kendi bahçenizde gördüğünüzde, size çukurun üzerinden atlayarak geldiği söylerse, ona inanabilirsiniz. Komşunuz küçük bir çukurun üzerinden rahatlıkla atlayabilir. Ancak çukurun genişliği 3 metre olsaydı komşunuzdan şüphe duyardınız.
Çukurun genişliği 100 metre olurda komşunuz size "üzerinden atlayarak geldim"
şeklinde bir cevap verirse, komşunuzun kesinlikle kuyruklu bir yalan attığından
emin olabilirsiniz.
Ancak komşunuz size "binlerce yıllık bir zaman içerisinde o devasal çukurun içerisinde tesadüflerle tepecikler oluştu ve ben oluşan her tepeciğin üzerinden atlaya atlaya buraya kadar geldim" şeklinde
bir cevap verirse söylediklerinin saçmalığı bir yana yalan olduğunu nasıl ispat
edebilirsiniz?
Elbette edemezsiniz.
Bu sayfada "Kısa gagalı İspinoz" ile "Uzun gagalı
İspinoz" kuşunun görünürde birbirinden evrimleştiği izlenimini verdiğini, ancak
moleküler düzeye indikçe değil 100 metrelik bir çukurun, genişliği kilometreleri
bulan kalyonların, vadilerin varlığına şahit olacağız.
- GERÇEĞİ GÖRMEK -
Gözün harikulade yapısı milyarlarca yıllık sayısız aşamalar neticesinde meydana gelmiş olabilir mi?
Darwin'e göre, görme olayı karanlık bir noktaydı. Ama içerisinde bulunduğumuz yüzyılda biyokimyacıların ortak ve yoğun çalışmalarından sonra, görme ile ilgili soruların cevaplarını hemen hemen çözmüş bulunuyoruz.
Aşağıdaki paragraf, gözün çalışmasının biyokimyasal bir özeti niteliğindedir.
İlginç isimler nedeniyle okuma azminizi kaybetmemelisiniz. Bunlar sadece
isimlerdir, arabalarla ilgili bir kitabı ilk defa olarak okuyan kişinin
karbüratör veya diferansiyel sözcükleri ile ilk kez karşılaşmasından pek farklı
değildir.
Hazır olun ! Az sonra bilgisayarınızın ekranından çıkacak olan bir foton, gözünüzün merceğinden geçerek retinaya çarpacak. O andan sonra neler olacağını merak ediyorsanız cevabı aşağıda.
. .
***Akademik
Bilgi***: Işık ilk olarak retinaya çarptığında, foton 11 cis retinal adı verilen bir molekül ile etkileşir. Bu molekül birkaç pikosaniye
içinde "trans-retina" denilen bölgeyi düzenleyebilir. Retina molekülünün şeklindeki değişiklik, retinanın sıkıca bağlandığı rodopsin
denilen proteinin şeklinde değişikliğe neden olur.
Proteindeki değişiklik, davranışlarını da değiştirmektedir. Bundan sonra metarodopsin
2, "transdusin" denilen başka bir proteine yapışır. Metarodopsin 2'ye çarpmadan önce transdusin, GDP denilen küçük bir moleküle sıkıca bağlanır. Ancak transdusin, metarodopsin 2 ile temasa geçtiğinde, GDP düşer ve GTP adı verilen başka bir molekül transdusine bağlanır. (GTP ve GDP birbirleriyle ilişkili, ama çok farklıdırlar) GTP transdusin metarodopsin 2 üçlüsü, artık fosfodiesteraz adı verilen başka bir proteine bağlanır.
Bu protein, hücrenin en iç kısmındaki zarına yerleşmiştir. Fosfodiesteraz proteini metarodapsin 2 ve diğerlerine bağlandığında, bir molekülü kesebilecek kimyasal bir yetenek elde eder ve cGMP denilen (GDP ve GTP'nin kimyasal akrabası) bir molekülü keser. Hücrenin içinde pek çok cGMP molekülü bulunmaktadır. Fakat fosfodiesteraz bunun derişimini
düşürür.
Bu aynı çekilen tıpanın, küvetin içindeki su seviyesini düşürmesine benzer. cGMP'yi bağlayan bir başka zar proteini de iyon kanalıdır. Hücredeki sodyum iyonlarını düzenlemek için bir giriş kapısı görevini görür. Normal olarak iyon kanalı, sodyum iyonlarının ayrı bir protein onları aktif olarak tekrar dışarı atarken, onların hücrede dolaşmalarını sağlar.
İyon kanalının bu iki yönlü hareketindeki, pompalama etkisiyle hücredeki sodyum
iyonları belirli bir seviyede kalır. Fosfodiesteraz proteinin faaliyetleriyle cGMP'nin miktarı azaldığında, iyon kanalları kapanır ve bu durum pozitif yüklü sodyum iyonlarının hücresel yoğunluğunun düşmesine neden olur. Sonuçta hücre zarındaki elektrik yükleri dengesizleşir ve en son aşamada bir elektrik akımının optik sinirlerden beyine doğru iletilmesine neden olur.
İşte bu son noktada beyin, iletilen elektrik akımını yorumlayarak görme olayı
gerçekleştirir.
Eğer yukarıda belirtilen reaksiyonlar hücrenin içinde gerçekleşen tek reaksiyonlar olsaydı, 11 cis retinal, cGMP ve sodyum iyonlarının kaynağı hemen tükenebilirdi. Harekete geçmiş olan proteinleri durduracak ve hücreyi eski şekline dönüştürecek bir şey gerekmektedir. Bunu birçok mekanizma yapar. Öncelikle karanlıkta iyon kanalı (sodyum iyonları ile birlikte) kalsiyum iyonlarının hücreye girmesini sağlar.
Kalsiyum, sürekli bir kalsiyum yoğunluğunun elde edilebilmesi için farklı bir
protein tarafından dışarı atılır. cGMP seviyesi düştüğünde, iyon kanalı kapanır
ve kalsiyum iyon yoğunluğu da azalır. cGMP'yi ortadan kaldıran fosfodiesteraz enzimi daha düşük kalsiyum yoğunluğunda yavaşlar. İkinci olarak, guanilat siklaz adı verilen bir diğer protein, kalsiyum seviyesi düşmeye başladığında cGMP'yi tekrar sentezlemeye başlar. Üçüncüsü, bütün bunlar olurken metarodopsin 2, rodopsin kinaz adı verilen bir enzim tarafından yenilenir. Yenilenmiş rodopsin
daha sonra "Arrestin "denilen başka bir proteine bağlanır. Bu da rodopsinin daha fazla transdusin molekülünü harekete geçirmesini engeller.
Böylece hücre, tek bir foton sinyali ile başlatılan etkileri sınırlandıracak
mekanizmaları da içermektedir.
Retinayı geçen ışık sonunda rodopsine düşer ve bundan sonra 11 cis-retinale yeniden dönüştürülmelidir ve tekrar başka bir görme işlemi için başlangıç noktasına geri dönmek amacıyla rodopsin tarafından bağlanır. Bunu başarmak için, trans-retinal kimyasal olarak bir enzim tarafından trans-retinol'a dönüştürülür. Yapıda iki hidrojen atomu daha vardır. İkinci bir enzim daha sonra molekülü ll cis retinale çevirir. Son olarak da, üçüncü bir enzim ll cis
retinale daha önceden eklenmiş olan hidrojen atomlarını kaldırır ve tam bir
döngü tamamlanmıştır.
Gördüğünüz gibi gözümüz gerçektende hayali bile kurulamayacak derecede karmaşık kimyasal reaksiyonlar neticesinde görme işlemini gerçekleştirir. Gözün içerisinde reaksiyona giren ve beyine impuls gönderilmesini sağlayan onlarca enzim vardır.
Eğer bu enzimlerden birisinin olmaması halinde impuls meydana getirilemeyecek ve
beyinde görüntü oluşamayacaktır.
İndirgenemez karmaşıklıktaki bu reaksiyonlar, göz için "Ya tamamıyla var olmalı
yada hiç olmamalı" ilkesini ortaya koyar.
Alıntılar; "Darwin'in on dokuzuncu yüzyılda evrimin açıklayamadığını söylediği görme olayı ve gözün anatomik yapısı, gerçekten de evrimci bir mantıkla açıklanamaz. Gözün aşamalı olarak meydana geldiğini iddia etmek için ortaya atılan senaryolar, "stereo sistem" nasıl meydana gelmiştir sorusuna "amplifikatöre hoparlörleri takarak, CD çaları, radyoyu ve teybi de ekleyerek" cevabını vermek gibidir. Darwin'in evrim teorisi, hoparlörlerin ve amplifikatörün kurulma sistemini ya açıklamalı veya bunu denememelidir bile."
[Behe. J. Michael / Darwin'in Kara Kutusu / syf. 31]
"Elbette evrim her zaman aşamalı olarak gelişmez. Fakat, göz gibi komplike, görünür şekilde tasarlanmış objelerin meydana gelişinde aşamalı bir evrim süreci olmalıdır. Eğer bu durumlarda da aşamalı olarak gerçekleşmezse o zaman evrimin açıklayıcı bir gücü kalmaz. Eğer aşamalar yoksa mucize olması muhtemeldir, buda yine bir açıklama olmadığını gösterir."
[River Out Of Eden / R. Dawkins / syf. 83]
Geroge Miklos, "Metazoaların Evrimi Esnasında Kompleks Organların
Ortaya Çıkışı / (1993)" kitabında göz ve göz benzeri kompleks organların
aşamalı olarak meydana gelmesi hususunda şunları söyler;
"O halde her tarafımızı kuşatmış bu evrim teorisi neyi tahmin ediyor?
Rastgele mutasyon, seleksiyon katsayısı gibi bir avuç dolusu varsayım öne
sürerek, zaman içinde gen frekanslarındaki değişikliklerini mi inceliyor?. Bu büyük evrim teorisinin içeriği gerçekten bu mu?"
Çukuru atlayarak geçtiğini söyleyen Charles Darwin'in göz konusunda şunları
söyler;
"Gözün odağını farklı uzaklıklara uydurması, içeri bırakılacak ışık tutarını ayarlaması, küresel ve renksel sapmayı (Abberation) düzeltmesi gibi eşsiz düzenlenişlerinin tümünün doğal seçmeyle oluşabildiğinin pek saçma göründüğünü açık yürekle itiraf ederim."
[Charles Darwin/Türlerin kökeni/s. 145/[Ocak 1990,Onur yayıncılık]
Bu noktada hemen şunu belirtmek gerekir ki ilkel canlıların göz yapıları net olarak görme işlemini yerine getirmez, objeleri bulanık olarak görür. Bu durum akılda, gözün aşamalı evrimi neticesinde en son mükemmel halini aldığını çağrıştırsa da o canlıların gözleri de tıpkı insan gözü gibi olması gerektiği gibi canlının ihtiyaç duyduğu fonksiyonlarını tam olarak yerine getirir.
İlkel canlılarında gözlerinde eksiltilemez karmaşık reaksiyonlar cereyan ediyor
olup aşamalı olarak meydana gelmesi veyahut daha üstün gözlere evrimleşmesi
moleküler düzeyde tam anlamıyla imkansızdır.
Richard Dawkins koyu bir evrimci olmasına karşın göz gibi aşırı kompleks bir organın ya aşamalı olarak evrimleşmek zorunda olduğunu yada mucizevi bir şekilde ortaya çıkması gerektiğini şöyle dile getiriyor;
GÖZ KAMAŞTIRAN HAREKET MEKANİZMASI
Sitemizin "ilginç canlılar" sayfasında bir bakterinin kamçısının kesitine yer vermiştik. Aşağıda ise bu şeklin ayrıntılı bir modelini göreceksiniz.
Bu modelin aşamalı evrim süreciyle meydana gelip gelemeyeceğini irdeleyeceğiz.
Şimdi şeklimizi görelim:
Böyle bir prensiple çalışan bir motorun çok karmaşık bir yapıda olması gerekir ve araştırılacak birçok yönü olduğu da kesindir. Motor için farklı modeller önerilmiş olsa da, bu modellerden hiçbiri aşamalı evrim süreciyle meydana gelmesine olanak verecek kadar basit bir yapıya sahip değildir. Bakteri kamçısı bir pervane sistemi kullanır. Bu nedenle diğer yüzme sistemlerinin gereksinimlerini karşılamalıdır. Bakteri kamçısının en az üç parçadan oluşması gerektiğinden (kürek, rotor ve motor) eksiltilemez bir karmaşıklığı vardır.
Bu nedenle kamçının aşamalı bir evrim sürecinden geçmiş olması imkansızdır.
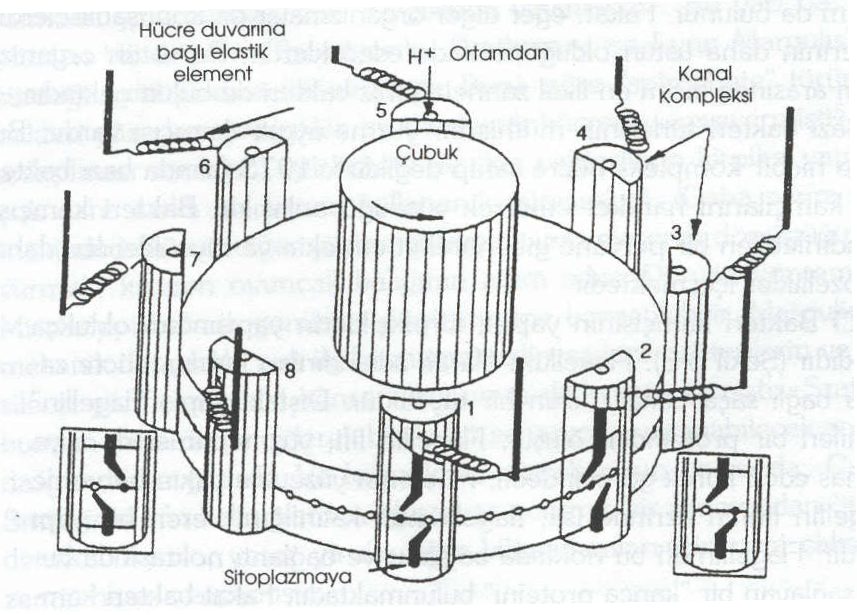
Yapılan deneyler, hareket mekanizmasının kaynağının kamçının tabanında yer aldığını göstermiştir. Elektron mikroskopu altında çeşitli halkamsı yapıların varolduğu gözlenmiştir. Kamçının döngüsel hareketlerinin kaçınılmaz sonuçları vardır. Bakteri kamçısının dönme hareketi aslında biyokimyasılar için çok şaşırtıcı ve beklenmedik bir buluştur. Hatta bu organeller fizik ile uğraşan bilim adamlarının bile dikkatinden kaçmamıştır. Mekanik hareketler oluşturan diğer sistemlerin aksine (örneğin kaslar) bakteriyel motor, hücre içinde ATP gibi bir molekülde saklı hazır enerjiyi kullanmaz.
Bunun yerine bakteri, zarından gelen bir asit akışından aldığı enerjiyi
kullanır.
***Akademik
Bilgi***: Bakteri kamçısı yaklaşık olarak 210 parça proteinden meydana gelmiştir, ancak bu yapının içerdiği karmaşıklık bizim tahminimizin çok üzerindedir. Bu proteinleri alıp böyle bir yapıya sahip olmayan bir hücreye enjekte ettiğinizde, bunlar faaliyet gösterebilen bir hareket oraneline dönüşmezler.
Bir hücrenin kirpikçiklere sahip olması için çok daha fazlası gerekmektedir.
Bu karmaşıklığın tüm sebepleri henüz bilinmemektedir ve daha fazla araştırma gerektirmektedir. Diğer proteinlerin bulunamamasının nedenleri arasında kirpikçiğin hücre içinde bağlanacağı başka yapıların da olması, kirpikçiğin elastikiyetinin değişmesi, çarpma hareketinin zamanlamasının değişmesi ve tüycüğe ait zarın güçlendirilmesi bulunmaktadır. Bakteri kırbacı (flagellum) ise yukarıda söz edilen proteinlerin yanısıra, fonksiyonlarını gerçekleştirebilmek için 40 kadar başka proteine ihtiyaç duymaktadır. Bu proteinlerin görevlerinin tam olarak bilinmemesine rağmen, motoru kapatıp açacak sinyalleri gönderirler.
Kırbacın hücre duvarına veya zarına saplayacak proteinleri oynatırlar, yapının
oluşmasında rol oynayan proteinlere yardımcı olurlar ve flagellumu oluşturan
proteinlerin düzenlenmesini sağlarlar.
Kısaca, biyokimyacılar kirpikçik ve kırbaç gibi görünürde basit olan yapıları incelemeye başladıkça, inanılmaz derecede bir karmaşıklıkla karşılaşmışlardır. Bunlar düzinelerce ve hatta yüzlerce ayrı parçadan oluşmaktadır. Aslında bizim burada üzerinden bile geçmediğimiz parçalar, kirpikçiklerin çalışabilmesi için gereklidir.
Gerekli parçacıkların sayısı arttıkça, sistemin bir araya getirilmesindeki
zorluk da artar ve ortaya atılan dolaylı senaryolar da çıkmaza girer.
Komşumuz Darwin de giderek daha çok hata yapmaya başlar. İlgili proteinler üzerinde yapılan çalışmalar, sistemin karmaşıklığını açıklamaya yetmemiştir. Problemin hassasiyeti çözümlenememiş, hatta giderek daha da içinden çıkılmaz bir hal almıştır. Evrim teorisi, kirpikçik veya kırbaç hakkında hiç bir açıklama yapamaz.
Yüzme sistemlerindeki bu aşırı derecedeki karmaşıklık, evrim teorisinin aslında
hiçbir zaman bir açıklama yapamayacağını da göstermektedir.
Evrimci yaklaşımla aşamalı bir gelişmeye ters düşen sistemlerin sayısı arttıkça, yeni bir açıklamaya ihtiyaç duyulmaktadır.
Kirpikçik ve bakteri kırbacı, Darwin'e problem çıkaran sistemlerin sadece
birkaçıdır. ***Akademik
Bilgi Sonu***
"Bakteri kamçısı hakkında yayımlanmış profesyonel eserlerin geçmiş yıllar içinde sayıları binleri aşmıştır ve kirpikçiklerle ilgili konular oldukça zengindir. Aslında bu şaşırtıcı değildir, çünkü kamçı müthiş bir biyofiziksel olaydır ve aynı zamanda tıp biliminde de önem taşımaktadır. Fakat yine burada da, evrimsel açıklamaların yetersizliği göze çarpmaktadır.
Bize tüm biyolojinin bir evrim gözlüğüyle incelenmesi gerektiği söylense de,
hiçbir bilim adamı bu olağanüstü moleküler makinenin nasıl evrimleştiği ile
ilgili bir model ortaya atamamıştır." [Behe. J. Michael/Darwinin Kara Kutusu/ syf.79]
Bakteri kamçıları ile ilgili meşhur bir model vardır. Bu modele göre kamçılı bir bakteri, tek hücreli bir Amip tarafından fagosite edilir (içeri alınır).
Daha sonra aşamalı bir evrim süreci içerisinde bakterinin kamçısı Amip'in kirpik
organına dönüşmüş, ve bu organel milyarlarca yıl korunarak insan ve diğer memeli
hayvanların soluk borusundaki silleri meydana getirmiştir.
Cevaplanması gereken birinci soru, bakterinin ilk var olduğu zamanlarda nasıl olup ta kompleks bir kamçıya sahip olduğudur. Bakteri kamçısının aşama aşama meydana gelmesi gerçektende olağan dışıdır.
Bunun imkansızlığını şekle bakarak ve açıklamaları okuyarak okuyarak sizde
görebilirsiniz.
İkinci problem ise kamçının sil gibi (her ne kadar sil ve kamçı birbirine benzese de) aşırı derece kompleks bir organele nasıl bir mekanizmayla evrimleştiğidir. Bir bakteri kamçısının bir sile dönüşmesi, bir "Otomobilin"in "Kamyon"a dönüşmesini
iddia etmekten farksızdır. Çünkü sil ve bakteri kamçısı moleküler düzeyde birbirlerinden oldukça farklı yapılara sahiptir.
Bu farklı organellerin aşamalı olarak evrimi ise moleküler düzeyde resmen
imkansızdır.
Bakteri kamçısı enerji kaynağı olarak H atomlarını (Otomobilin benzini yakması) kullanırken, siller ise enerji kaynağı olarak ATP'yi (Kamyonun mazotu kullanması) kullanmaktadırlar. O halde bir kamçıyı bir sile aşamalarla evrimleştirmek, ancak bilinçli bir müdahele neticesinde gerçekleştirilebilir ki doğada böyle bir mekanizma yoktur.
Bu mekanizmanın mutasyonlar olmasını düşünmek ise tam manasıyla imkansızdır.
Kısacası otomobili kendi halinde tesadüflerle bir Kamyona
evrimleştiremezsiniz.
Alıntı; "Flagellum olarak adlandırılan bakteri kamçısı, hücre zarına bağlı kıl'a benzer uzun bir tüycüktür. Dıştaki yapı, "flagelİin" denilen bir proteinden oluşur. Flagellin lifi, yüzme sırasında suya temas eden kürek görevindedir. Hücrenin yüzeyine yakın bir yerde flagellin lifinin üzrinde ise, flagellumun kalınlığını veren bir yapı vardır.
Flagellin lifi bu noktada bağlanır ve bağlantı noktasında bunu sağlayan bir
"kanca proteini" bulunmaktadır.
Fakat bakteri kamçısının, sillere benzer bir motor özelİiği bulunmamaktadır. Yani bakteriden koptuğunda, hareketsiz sitoplazma içerisinde kalır. Bu nedenle hareketli kamçıyı sillerdekine benzer bir mekanizmaya aşamalı olarak evrimleştirmek açıkça imkansızdır."
[Behe. J. Michael. Darwinin Kara Kutusu/s. 78]
PARMAĞIMIZA BATAN
DİKEN
Elinize bir iğne alıp parmağınıza batırın ve damarlarınızdan akan kanın bir süre sonra pıhtılaştığını seyredin. Bu pıhtılaşma size oldukça sıradan gelir.
Kan sürekli olarak akar ve nihayetinde katı bir hal alarak iğnenin açtığı deliği
tıpa gibi kapar.
Charles Darwin de Galapagos adalarında gezerken eline batan diken yüzünden akan kanın ne gibi işlemler neticesinde pıhtılaştığını bilmiyordu. Bu yüzden
"Türlerin Kökeni" kitabını dar bir bilgi çerçevesinde yayınladı.
Kan pıhtılaşması için "Uçuşa geçmek" deyimi oldukça yerinde olur. Çünkü
parmağınızdan akan kanda meydana gelen reaksiyonlar, olayın tahmininizden ne
kadar karmaşık olduğunu gözler önüne sermektedir.
***Akademik
Bilgi***: Vücut genellikle daha sonra kullanmak üzere aktif olmayan enzimleri depolar (enzimler kimyasal bir reaksiyonu harekete geçiren proteinlerdir, fibrinojenin kesilmesinde olduğu gibi). Bu aktif olmayan enzimlere, proenzim denir. Belirli bir enzimin gerekli olduğuna dair bir sinyal alındığında, ilgili proenzim harekete geçerek gerçek bir enzim oluşturur.
Fibrinojenin fibrine dönüşmesi gibi, proenzimler de kendi üzerinde belirli bir
noktadan bir parçanın kesilmesiyle olgun bir enzime dönüşürler.
Bu strateji genellikle sindirimle ilgili enzimlerde kullanılmaktadır. Büyük miktarda aktif olmayan enzim depolanır ve bir sonraki öğünün gelişiyle hemen harekete geçer. Trombin genelikle aktif olmayan, protrombin halinde mevcuttur. Aktif olmadığı için protrombin, fibrinojeni kesemez ve böylece canlı, kontrolsüz bir pıhtılaşmanın ölümcül etkilerinden korunmuş olur. Fakat buradaki kontrolün sırrı hala çözülememiştir. Fibrinojen ve protrombin, kanın pıhtılaşması sisteminde kullanılan proteinlerin tamamı olsaydı canlının durumu herhalde pek iyi olmazdı.
Hayvan yara aldığında amaçsızca dolaşan protrombin fibrinojenin yanından geçip
gidecek ve canlı kan kaybından ölecekti.
Protrombin fibrinojeni keserek fibrine dönüştüremediğinden, protrombini harekete geçirecek bir mekanizmaya ihtiyaç vardır. Bu sistemde. Bir parça diğerini harekete geçirir ve bu böyle devam eder. Stuart faktörü denilen bir protein de protrombini keser ve onu aktif trombine dönüştürür. Ancak bu şekilde trombin, fibrinojeni fibrine dönüştürür ve kan pıhtısını oluşturur. Fakat ne yazık ki, Stuart faktörü ve protrombin ile fibrinojen, kanın pıhtılaşmasında rol oynayan tek proteinler olsaydı, Stuart faktörü şelale etkisini hemen başlatacak ve organizmanın kanını kurutacaktı. Bu nedenle Stuart faktörü de kanda aktif durumda bulunmamaktadır ve harekete geçmesi için aktifleştirilmesi gerekmektedir.
Bu noktada aklımızı karıştıran bir yumurta-tavuk senaryosuyla karşı karşıya
kalırız.
Aktif durumdaki Stuart faktörü bile protrombini harekete geçirmeye yeterli değildir. Stuart faktörü ve protrombini bir test tübüne koyup karıştırabilirsiniz, fakat bu sırada trombin oluşana kadar canlı kanamadan ölüp gidecektir. O halde görülmektedir ki, akselerin adında başka bir protein Stuart faktörünün harekete geçmesi için gereklidir.
Dinamik ikili akselerin ve aktif Stuart faktörü, protrombini hemen keser ve
hayvanın kanaması durdurulur.
Yani, bu aşamada bir proenzimi aktifleştirebilmek için, iki ayrı proteine ihtiyaç duyulmaktadır. Akselerin de başlangıçta aktif olmayan proakselerin durumundadırlar.
(derin bir nefes alabilirsiniz).
Peki onu ne aktifleştirir? Trombin! Fakat trombin hatırlayacağmız gibi bu şelalede, proakselerinin durduğu yerden daha aşağıdadır. Bu durumda akselerin üretiminde rol oynayan trombin, torunun anneannenin doğumundan önce varolmasına benzer. Ne var ki, Stuart faktörünün protrombini çok yavaş bir hızda kesmesi nedeniyle, kanda her zaman bir miktar trombin bulunmaktadır.
Kanın pıhtılaşması bu nedenle otomatik-kataliz özelliğine sahiptir, çünkü
şelaledeki proteinler aynı proteinlerden daha fazla üretilmesini de sağlar.
Burada biraz geriye gitmemiz gerekir, çünkü hücrede oluşturulan protrombin, aktif durumdaki Stuart faktörü ve akselerin olmasına rağmen trombine dönüşmemektedir. Protrombin öncelikle değiştirilmelidir ve bunu yaparken on çeşit aminoasit artık oluşturur. Glutamat (Glu) maddesi, Karboksiglutamat (Gla) maddesine dönüşür. Bu değişim, alt çeneye bir üst çene eklemeye benzer. Tamamlanan yapı ısırabilir ve dişlerin arasında bir nesneyi tutabilir, fakat alt çene olmadan bu yapılamazdı. Protrombinde ise Gla artıkları kalsiyumu ısırır (bağlanır) ve böylece protrombin hücrelerin dış yüzeylerine yapışabilir.
Sadece bu değişime uğramış kalsiyum-protrombin ikilisi hücre zarına bağlanmış
durumuyla, Stuart faktörü ve akselerin tarafından aktifleştirilerek trombine
dönüştürülebilir.
Protrombinin değiştirilmesi bir kaza eseri değildir. Tüm biyokimyasal reaksiyonlarda olduğu gibi bunu yapacak özel bir enzime ihtiyaç vardır. Bu enzimin yanısıra, Glu maddesinin Gla'ya çevrilebilmesi için de K vitamini gereklidir. K vitamini bir protein değildir, fakat görme için gerekli olan 11 cis retinal (gözdeki protein gibi) küçük bir moleküldür.
Mermilere gereksinimi olan bir silah gibi, Glu'yu Gla'ya dönüştüren enzimin de K
vitaminine ihtiyacı vardır.
Hala pek fazla gelişme kaydettiğimiz söylenemez. Şimdi geriye dönüp Stuart faktörünü neyin harekete geçirdiğini tekrar görmeye çalışalım. Bunun iki farklı yoldan başarılabilmesi mümkündür, iç ve dış yöntemlerle. İçteki yöntemde pıhtılaşma için gerekli olan tüm proteinler kan plazmasında yer alır, dıştaki yöntemde ise bazı pıhtılaşma proteinleri hücrelerin içindedir.
Şimdi ilk olarak iç yöntemi inceleyelim.
Bir hayvan yaralandığında, yaranın yakınındaki hücrelerin yüzeyinde "Hageman" faktörü denilen bir protein açığa çıkar. Hageman faktörü daha sonra HMK adlı başka bir protein tarafından kesilir ve aktif duruma gelir. Aktifleşen Hageman faktörü, başka bir protein olan "Prekallikrein"i aktif hale sokar ve "Kallikrein"e dönüştürür. Kallikreinin varlığı ise, HMK'yı hızlandırarak daha çok Hageman faktörünün aktifleştirilmesini sağlar. Aktif Hageman faktörü ve HMK, birlikte konvertin proteinini harekete geçirerek, Christmas faktörü denilen proteini de aktifleştirir. Sonuç olarak, aktif durumdaki Christmas faktörü ve Antihemofili faktörü, Stuart faktörünü aktif haline getirir. İçteki yöntem gibi, dıştaki yöntem de bir şelaleyi andırmaktadır. Dış yöntem ise "Prokonvertin"denilen proteinin, aktif Hageman faktörü ve trombin ile birlikte konvertine dönüştürülmesiyle başlar.
Başka bir proteinin varlığında ise, doku faktörü gibi, konvertin Stuart
faktörünü aktif forma sokar.
Doku faktörü genellikle hücrelerin kanla temas etmeyen dış kısımlarında bulunmaktadır. Bu nedenle yalnız yaralanma durumlarında doku kanla temas eder ve dış yöntem uygulamaya geçer. İç ve dış yöntemler çeşitli noktalarda kesişirler. İç yöntemle aktifleşen Hageman faktörü, dış yöntemdeki prokonvertini çalıştırabilir. Böylece konvertin iç yöntemi takip ederek, aktif PTA ' nın Christmas faktörünü aktifleştirir. T rombin ise iki yöntemi de harekete geçirebilmektedir, çünkü pıhtılaşma şelalesindeki Antihemofili faktörünü aktifleştirebilir.
Bu durumda Christmas faktörü, Stuart faktörünü dönüştürebilir ve onu aktif hale
sokar, ayrıca prokonvertini de aktifleştirebilir.
***Akademik
Bilgi Sonu***
Buna bir örnek olarak Hemofili hastalarını verebiliriz. Hemofili hastalarının Faktör-8 proteinini şifreleyen gende mutasyonları olduğu için üretimi yapılamaz, ve hastanın kanı pıhtılaşma gösteremez.
Bu durum kişinin en ufak bir yaralanmada kan kaybından ölümüne yol açar.
Ancak daha macera sona ermedi. Kanın pıhtılaşmasının ardından belli bir noktada durdurulması gerekir.
Çünkü pıhtılaşma durdurulmaz ise kanınızın son damlasına kadar pıhtılaşma ilerleyecek ve hayatınızı kaybedecektiniz.
Bu sistemin kontrol mekanizması yukarıda anlatılan kadar karmaşık bir dizi
reaksiyonu da beraberinde getirir.
Cesaretiniz kırılmadıysa okumaya devam edebilirsiniz.
Pıhtılaşmanın Kontrol Altına Alınması:
Öncelikle, antitrombin denilen plazma proteini aktif pıhtılaşma proteinlerine bağlanır ve onları aktif olmayacakları hale getirir. Eğer "Heparin" adı verilen bir maddeye bağlanmazsa, antitrombin de aktif değildir. Heparin zarar görmemiş kan hücrelerinin ve damarların içinde oluşur. Pıhtılaşmanın lokalize olduğu ikinci yol C proteinin hareketleri doğrultusundadır. Trombin tarafından aktif hale getirildikten sonra C proteini, akselerini ve aktif antihemofili faktörünü yok eder. Sonuçta, "Trombomodulin"adı verilen bir protein kan damarlarının içindeki hücrelerin yüzeyinde sıralanır. Trombomodulin trombine bağlanır, trombinin fibrinojeni kesme kabiliyetini azaltır ve aynı zamanda C proteinini aktif hale getirme kabiliyetini arttırır.
Pıhtı ilk olarak oluştuğunda, oldukça hassastır.
Eğer yara alan bölge bir darbe alırsa, pıhtı kolayca zarar görebilir ve kanama tekrar başlar. Bunu önlemek için, vücudun pıhtıyı güçlendirme metodu vardır. Birikmiş fibrin, aktif hale getirilmiş bir protein olan FSF "fibrin stabilizasyon faktörü"(fibrin sabitleme faktörü) tarafından birbirine bağlanır. FSF, farklı fibrin molekülleri arasında karşılıklı kimyasal bağlar meydana getirir. Sonuçta yara iyileştikten sonra pıhtı ortadan kaldırılmalıdır. Plazmin denilen bir protein, özellikle fibrin pıhtılarını kesmek için bir makas görevi görür. Neyse ki, plazmin fibrinojen üzerinde etkin değildir. Ancak plazmin çok hızlı hareket edemez, fakat bu sayede yaranın tamamen iyileşmesi için zamanı olur. Bu nedenle "Plazminojen" adı verilen aktif olmayan bir form içinde meydana gelir. Plazminojenin plazmine dönüşümü ise TPA adı verilen bir protein tarafından sağlanır. Pıhtının çözülmesini kontrol eden başka proteinler de vardır. Bunların arasında, plazmine bağlanan ve fibrin pıhtılarını bozmasını engelleyen A2-antiplazmin bulunmaktadır.
***Akademik
Bilgi Sonu***
Bu maceraların anlamı, sizin gençlik zamanlarınızda top oynarken dizinizi
yaralamanıza rağmen hayatta kalmanızda yatar.
Bu hiper-kompleks sistemin aşamalı bir rastlantısal mutasyonlar sonucu
meydana gelmesi mümkün olabilir mi?
Amerika'da yayınlanan "Trombosis and Haemostatis" isimli bir dergide Profesör Russel Doolittle, kanın pıhtılaşma reaksiyonları ile ilgili uzun araştırmalar sonucunda (daha doğrusu uzun bir düşünme sürecinden sonra) bir makale yayımlamıştır.
Kanın pıhtılaşmasıyla ilgili bir dizi senaryo üretmiş, ve nihayetinde makalede
şu ifadelere yer vermek zorunda kalmıştır.
''Bu kompleks ve hassasiyetle dengelenen süreç nasıl evrimleşmiş olabilir?
Paradoks burada yatıyor, eğer her protein bir başka proteinin aktivasyonuna
bağlı ise, bu sistem nasıl meydana gelmiştir? Bu düzen tamamıyla oluşmadan bu
sistemin parçaları ne işe yarayabilirdi?"
Biyokimya Prf. Michael Behe ise kanın pıhtılaşma senaryolarına ilişkin olarak
şunları söyler;
"Eğer pıhtılaşma reaksiyonlarını kontrol altına alan ikinci bir kontrol mekanizma olmasaydı, canlının kanı son damlasına kadar pıhtılaşacaktı. O halde kanın pıhtılaşma sisteminin, pıhtılaşmayı kontrol altına alan ikinci bir mekanizmayla aynı anda ortaya çıkmış olması gerekir. Bu yüzden dünyada hiç kimsenin "kan pıhtılaşması mekanizmasının evrimi" konusunda mutlak bir bilgisi yoktur."
Aşamalı bir evrim süreci ile meydana gelememesinin nedeni, zincirleme
reaksiyonları tetikleyen faktörlerin çok fazla sayıda olması ve her birinin
tesadüfi mutasyonlarla meydana gelme olasılığının çok düşük olmasıdır.
Pıhtılaşma reaksiyonlarının herhangi bir noktasına gidelim. Girdiğimiz noktada bir faktör olsun (örneğin Stuart faktörü). Söz konusu faktör önüne çıkan bir Protrombini keserek onu aktif hale yani Trombin'e çevirecektir. Trombin bu aşamadan sonra fibrinojeni keserek onu fibrin haline çevirir.
Fibrin artık aktiftir, fakat aşamalı evrim sırasında henüz ortalarda fibrinojen
yoksa trombin ne yapabilirdi?. Yapabileceği tek şey, fibrinojenin tesadüfen
mutasyonlarla meydana gelmesi için milyonarca yıl beklemesidir.
Ancak bir proteinin tesadüflerle meydana gelmesi neredeyse imkansızdır. Bir protein (burada proteinimiz fibrinojen) yaklaşık 105-1000 amino asitten meydana gelir. Ancak ben burada en az sayıda amino asit ihtiva edeni ele alacağım. 105 amino asitlik bir proteinin meydana gelme olasılığı, 20 üzeri 105 dir. Bu gerçektende çok küçük bir rakamdır. Bu proteinin tesadüfen meydana gelen bir mutasyonla meydana gelmesini beklemek için evrenin milyar yıllık yaşı bile yetmemektedir. Halbuki damarından kanı akan zavallı adamın beklemeye vakti yoktur !
Durum göründüğünden daha vahimdir. Bu olasılık, sözünü ettiğimiz onlarca faktör proteinlerinden yanlızca birine aittir. Her bir protein için aynı olasılık söz konusudur. Ancak her protein 104 aminoasitten oluşmaz !. Burada en büyük engeli ise Darwin'in doğal seleksiyonu oluşturur. Doğal seleksiyona göre canlının, işe yaramayan organ veya metabolik elemanları doğal seçme ile ayıklanır.
O halde bu adamın yaşaması imkansızdır, soyu tükenecektir.
Tek bir şart vardır o da, tesadüfen meydana gelen astronomik sayıda yararlı mutasyonlarla her bir faktörün aniden ortaya çıkmasıdır.
Bu ise tamamen hayal gücünün sınırları dışına çıkmak anlamına gelmektedir.
Alıntı;
Buraya kadar olan bölüm adeta insanın başını döndürmektedir. Her bir protein diğerinin aktif komplementeri
(tamamlayıcısı) gibi iş görür ve zincirleme reaksiyonun devamlılığını sağlar. Bu
faktörlerden birisinin olmaması sistemi çökertecek ve kanın pıhtılaşmasını
engelleyecektir.
***Akademik
Bilgi***: Pıhtılaşma bir kez başladığında, hayvanın
sahip olduğu kanın tamamı katılaşana kadar pıhtılaşmasını engelleyecek faktörler
nelerdir? Pıhtılaşma çeşitli nedenlerden dolayı sadece yaranın bulunduğu bölgeyi
kapsar.
Bütün bu maceraların anlamı nedir?
Alıntılar:
2. Bölüm İçin Tıklayınız
![]()
|
|
instagram.com/ahmet.eksik |
|
|
biyolojidunyasi@hotmail.com |